L'epitelio ciliato riveste la mucosa in tutte le varie porzioni delle vie respiratorie compresi i seni paranasali, la tuba uditiva e l'orecchio medio.
Attività ciliare e muco bronchiale costituiscono un'unità funzionale detta clearance muco-ciliare che contribuisce alle difese dell'apparato respiratorio in quanto meccanismo responsabile del normale trasporto del muco e della conseguente eliminazione dalle vie respiratorie di particelle di polvere, cellule morte, batteri e virus inalati ed in esso inglobati e quant'altro possa danneggiare l'apparato respiratorio. Condizioni essenziali per un'efficace clearance muco-ciliare sono la composizione ottimale del muco (glicoproteine, proteoglicani e lipidi) ed una valida attività ciliare. In condizioni normali il muco si organizza sulla superficie delle vie aeree in 2 strati a diversa viscosità, che vengono spostati verso l'orofaringe dal movimento delle ciglia presenti all'apice delle cellule dell'epitelio respiratorio.
Circa l'80% delle cellule presenti nell'epitelio respiratorio sono ciliate, mentre circa il 20% è rappresentato da cellule secernenti muco.
Ciascuna cellula ciliata presenta sulla sua superficie apicale circa 200-250 ciglia, il cui diametro è di circa 250 nm. La lunghezza delle ciglia varia da 5 a 8 μm riducendosi man mano che si procede verso la periferia dell'albero bronchiale. Le ciglia dell'epitelio respiratorio sono dotate di movimento, attraverso il quale provvedono al drenaggio verso l'esterno del muco presente nelle vie aeree e di norma battono ad una frequenza di circa 12 Hz, almeno a livello nasale.
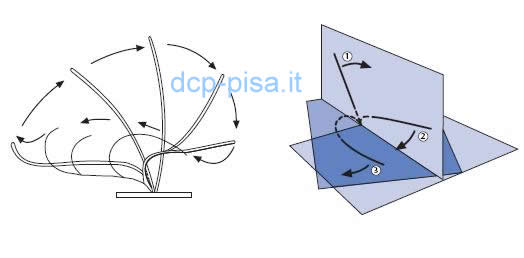
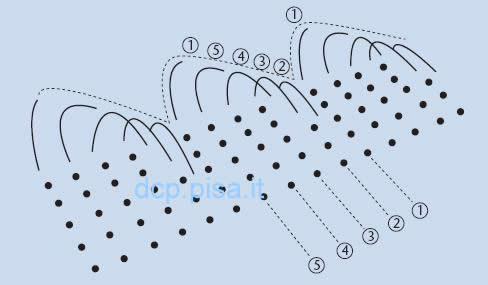
Fino a pochi anni fa si è ritenuto che il battito ciliare di norma fosse asimmetrico e caratterizzato da un movimento in avanti su un piano verticale, relativamente rigido, capace di produrre uno spostamento di muco, seguito da un movimento di ritorno con una flessione su un piano verticale ed orizzontale che non riporta il muco all'indietro. In realtà con l'aiuto di un nuovo videoregistratore ad alta velocità di immagini digitali ad elevata risoluzione che ha consentito di visualizzare il movimento ciliare in tre diversi piani è stato recentemente messo in evidenza che le ciglia battono semplicemente in avanti e indietro sullo stesso piano senza il rapido movimento di recupero sul piano obliquo.
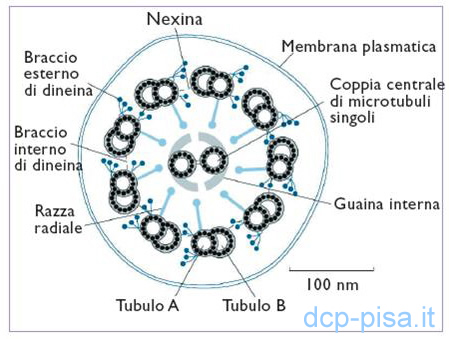
I microtubuli sono lunghi tubi cavi di natura proteica del diametro esterno di 25 nm. Sono formati da dimeri della tubulina, polipeptide presente in forma di subunità globulari (α e β), ciascuna composta da circa 450 aminoacidi. I dimeri di tubulina (α e β) allineati in successione formano dei protofilamenti i quali si affiancano e si dispongono intorno ad una cavità centrale dando origine ai microtubuli. Nei protofilamenti i dimeri α e β sono sfalsati tra loro in modo che la tubulina α di un dimero è in contatto con la tubulina β di quello vicino. Ogni coppia periferica di microtubuli è costituita da una subfibra A e da una subfibra B. La subfibra A è un microtubulo completo che in sezione trasversale è formato da 13 protofilamenti, la subfibra B è un microtubulo incompleto ed è formato da 11 protofilamenti. I microtubuli centrali sono anch'essi formati da 13 protofilamenti di tubulina.
Il processo che porta alla formazione dei microtubuli in vitro avviene spontaneamente in presenza di GTP, mentre in vivo esistono centri di organizzazione che forniscono una base da cui i microtubuli possono accrescersi: i corpuscoli basali, che agiscono come stampo per disporre i microtubuli a coppie con il pattern 9+2.
Il corpuscolo basale è una struttura cilindrica posta nella porzione apicale del citoplasma della cellula ciliata che contiene un anello di 9 triplette di microtubuli (A,B,C) e ogni tripletta contiene un microtubulo completo (A) fuso con altri due incompleti (B e C). I microtubuli periferici dell'assonema, si continuano in quelli del corpuscolo basale e a questo livello a ciascuna coppia si aggiunge un terzo microtubulo incompleto, mentre si ha l'arresto della coppia centrale 1µ sopra la superficie cellulare. Come nell'assonema, anche nel corpuscolo basale esistono legami proteici di stabilizzazione, in particolare tra la subfibra A di una tripletta e la subfibra C di quella vicina. Inoltre un “basal foot” si proietta dal corpuscolo basale nella direzione del battito ciliare.
L'assetto 9+2 dei microtubuli dell'assonema si modifica anche nella porzione apicale del ciglio dove le coppie periferiche perdono progressivamente un microtubulo (subunità B) ed altre strutture quali, ad esempio, la coppia centrale ed i bracci di dineina.
I microtubuli che compongono l'assonema vengono stabilizzati da proteine (MAPs: microtubule associated proteins), la più conosciuta delle quali è la dineina. Le molecole di dineina sono organizzate a formare coppie di bracci, interno ed esterno, che in senso antiorario, se osservati dalla base del ciglio, sporgono dalla subfibra A di una coppia alla subfibra B della coppia adiacente a intervalli regolari di 24 nm. Nell'assonema la dineina costituisce un grande complesso macromolecolare composto da 9-12 subunità di dimensioni diverse. Il braccio esterno, correlato con la frequenza del battito, è formato da 2 subunità α e β, catene pesanti (450.000-400.000 D) che possiedono attività ATPasica e formano le teste globulari del complesso, da 2 o più catene intermedie e da 4-8 subunità più piccole, le catene leggere che, determinano l'ancoraggio del complesso dineinico alla superficie del microtubulo della subfibra A. I bracci interni di dineina, che invece influenzerebbero la forma del battito, hanno una struttura di base essenzialmente simile a quella degli esterni, ma sembrano esistere in tre distinti tipi che contengono isomeri diversi di dineina.
Altre proteine accessorie sono la tectina simile ai filamenti intermedi, di 2-3 nm. di diametro che scorre internamente ai microtubuli tra le molecole di tubulina e sembra che contribuisca a formare la parete condivisa tra le subfibre A e B stabilizzandone la struttura; la nexina che, a intervalli di 86 nm, forma legami dalla subfibra A alla subfibra B delle coppie periferiche di microtubuli, simili a fasce intorno all'intero assonema paragonabili ai cerchi di un barile; i raggi o razze o ponti radiali che sporgono da ogni coppia con una periodicità di 29 nm. e terminano con la parte globulare vicino alla guaina interna, connettendo quindi ogni coppia periferica a quella centrale ed infine la guaina interna formata da sottili bracci proteici, le proiezioni, che sporgono con una periodicità di 14 nm dal paio di tubuli centrali e sono incurvati intorno ad essi quasi a formare un anello. Quest'ultima interviene, con i raggi e la nexina, nella regolazione della forma del battito ciliare.
Si è dimostrato sperimentalmente che il movimento ciliare non dipende dalla presenza del corpuscolo basale, né da quella della membrana plasmatica poiché l'assonema isolato è in grado di compiere movimenti di flessione se immerso in una soluzione salina contenente ATP e ioni magnesio oppure calcio. Inoltre esponendo assonemi isolati all'azione di enzimi proteolitici che distruggono nexina e ponti radiali, ma lasciano intatti microtubuli e dineina, in presenza di ATP (anche 10 µM) si osserva un allungamento dell'assonema fino a 9 volte la lunghezza iniziale. Infatti i microtubuli esterni scorrono l'uno sull'altro come gli elementi di un cannocchiale.
Sicuramente artefici del meccanismo di scorrimento sono quindi i bracci di dineina: sperimentalmente nell'assonema nudo la loro assenza provoca l'arresto del movimento. Questo ricompare con l'aggiunta di dineina e ATP. In assenza di ATP, i bracci di dineina non occupano tutto lo spazio presente tra una coppia e quella vicina e sono disposti a formare un angolo retto rispetto ai microtubuli. In presenza di ATP, invece, prendono contatto con la coppia adiacente e modificano il loro orientamento dirigendosi verso il basso in direzione del corpuscolo basale. Poiché la dineina è un' ATPasi, con l'idrolisi dell'ATP ogni braccio di dineina genera una forza che agisce in modo da spostare verso l'apice del ciglio la coppia di microtubuli adiacenti con cui prende contatto. Se tutti i bracci si attivassero nello stesso momento, l'assonema si avvolgerebbe a formare un'elica stretta, per cui è proprio la loro attivazione asincrona a permettere il tipico movimento a frusta del ciglio che è l'unico efficace dal punto di vista fisiologico. In presenza di nexina e di ponti radiali, che offrono punti fermi di ancoraggio, il movimento di scorrimento si trasforma in movimento di flessione.
Da studi effettuati su un'alga unicellulare provvista di 2 flagelli (Chlamidomonas) sembra che in condizioni di riposo l'attivazione dei bracci di dineina sia inibita da altre proteine e che il segnale trasmesso dai raggi e dalla guaina interna tolga questa inibizione promuovendo l'attivazione organizzata dei bracci di dineina lungo il flagello.